
"Биология. Человек. 9 класс". А.С. Батуев и др. (гдз)
Хромосомы. Клеточный цикл. Митоз растительной клетки
Таблица 5. Число хромосом соматических клеток некоторых видов организмов
| Виды | Число хромосом |
| Малярийный плазмодий Гидра Таракан Комнатная муха Сазан Окунь Зеленая лягушка Голубь Кролик Шимпанзе Человек |
2, 32 48 12 104 28 26 80 44 48 46 |
Таблица 6. Классификация хромосом по размеру и расположению центромеры
| Группа хромосом |
Номер по кариотипу |
Характеристика хромосом |
| A (I) B (II) C (III) D (IV) E (V) F (VI) G (VII) X-хромосома (относится к группе (III) Y-хромосома |
1, 2, 3 4,5 6—12 13—15 16—18 19—20 21—22 23 23 |
1 и 3 — почти метацентрические; 2 — крупная субметацентрическая Крупные субмета центрические Средние акроцентрические Средние акроцентрические Мелкие субметацентрические Самые мелкие метацентрические Самые мелкие акроцентрические Мелкая акроцентрическая |
Митоз растительной клетки
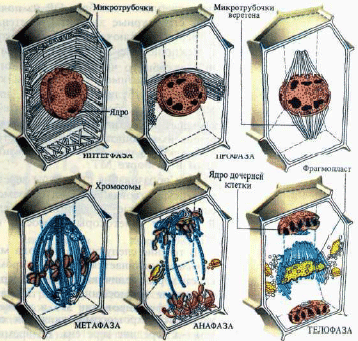
Рис. 23. Схема фаз митоза растительной клетки
В процессе митоза растительной клетки происходят сложные последовательные изменения структуры ядра и цитоплазмы, подразделяющиеся на фазы (рис. 23).
Первой фазой митоза является профаза — фаза реорганизации клетки. Разрушается ряд структур, существовавших в клетке до митоза, и строятся новые, связанные с процессом деления. В цитоплазме органоиды смещаются от ядра на периферию клетки. Ядро увеличивается в объеме, хроматин оформляется в хромосомы с кинетохором (определенный участок хромосомы) и с двумя сестринскими хроматидами каждая. Ядрышко постепенно диссоциирует. После распада оболочки ядра на ряд ретикулярных элементов в клетке формируется структура веретена.
В клетках растений нет центриолей (организаторов веретена), характерных для животных клеток. Их функцию выполняют скопления мембран ЭПС на полюсах клетки. С ними связаны структурные элементы веретена.
В течение следующего периода митоза — метафазы начинается движение хромосом. Перед его началом кинетохоры увеличиваются в размерах, от них отходят многочисленные хромосомные МТ. Полагают, что кинетохор участвует в формировании МТ и движениях хромосом. В течение митоза хромосомы движутся сначала к полюсам, а затем —к середине веретена. Во время этих перемещений в хромосоме раскручиваются две сестринские хроматиды, которые остаются соединенными в кинетохоре. Повреждение кинетохора в это время избирательным ультрафиолетовым облучением или лазером останавливает движение хромосом.
В результате перемещения хромосомы собираются вдоль поперечника веретена и образуют метафазную пластинку (метафаза). При этом они совершают небольшие перемещения вдоль веретена. В течение метафазы продолжается синтез РНК и белков, хотя и с невысокой скоростью.
Переход клетки к анафазе сопроиождается делением кинетохора, физическим разделением двух сестринских хроматид и перемещением разделившихся хромосом к полюсам кинетохором вперед. Происходит также перераспределение микротрубочек: количество их у полюсов уменьшается и увеличивается в интерполярной области, в районе эватора веретена. В поперечной плоскости веретена начинает образовываться зона скопления везикул — начало формирования разделительной пластинки.
После расхождения хромосом к полюсам начинается последняя стадия митоза — телофаза. У полюсов МТ веретена дезинтегрируют, образуются ядрышки, ядра, заканчивается формирование разделительной пластинки — фрагмопласта, делящей клетку пополам в экваториальной плоскости. В анафазе вдоль экватора веретена скапливаются пузырьки различного размера. Мелкие везикулы являются производными аппарата Гольджи содержат пектиновые вещества. В экваториальной плоскости обнаруживаются также мембраны ЭПС. Для образования фрагмопласта важно также присутствие кальция. Сливаясь, везикулы образуют две мембраны — плазмалеммы дочерних клеток, разделенные полужидким слоем, состоящим из пектиновых веществ. Взаимодействие везикул происходит между микротрубочками веретена.
От центра к периферии фрагмопласт растет за счет присоединения (самосборки) новых пузырьков, но цитоплазма дочерних клеток остается связанной через плазмодесмы, формирующиеся в тех участках клеточной пластинки, в которых сказались локализованными нити веретена с МТ и элементами ЭПС. Со стороны цитоплазмы дочерних клеток начинается формирование первичных клеточных стенок, а фрагмопласт превоащается в срединную пластинку, разделяющую материнскую клетку на две дочерние. Откладывающиеся микрофибриллы целлюлозы первичных клеточных стенок имеют рыхлую текстуру, но основное направление ориентации микрофибрилл перпендикулярно продольной оси клетки. Содержание целлюлозы в первичных клеточных стенках вначале не превышает 2,5%.
После окончания деления дочерние клетки растут благодаря синтезу компонентов цитоплазмы. Именно так растут и животные к метки. Как правило, дочерние клетки достигают размеров материнской и затем могут вновь перейти к делению. Процесс деления (митоз) и период цитоплазматического роста и подготовки к делению (интерфаза) составляют митотический цикл клетки.
Клеточный цикл
Один из постулатов клеточной теории гласит, что увеличение числа клеток, их размножение, происходит путем деления исходной клетки. Это положение полностью исключает какое-либо «самозарождение» клеток или их образование из неклеточного «живого вещества». Обычно делению клеток предшествует редупликация их хромосомного аппарата, синтез ДНК. Это правило является общим для прокариотических и эукариотических клеток.
Время существования клетки как таковой — от деления до деления — обычно называют клеточным циклом. Величина его может быть различной для разных типов клеток. Так, например, для бактериальных клеток в стационарных условиях культивирования это время может быть равно 20—30 мин. У эукариотических одноклеточных организмов время жизни клетки, продолжительность ее клеточного цикла, значительно больше. Так, инфузория туфелька может делиться 1—2 раза в сутки, время клеточного цикла при бесполом размножении у амёбы составляет около 1,5 суток, у инфузории трубача — 2—3 суток. Время прохождения клеточного цикла зависит от температуры и условий окружающей среды.
Клетки многоклеточных организмов обладают разной способностью к делению. Если в раннем эмбриогенезе клетки животных организмов делятся часто, то во взрослом организме они большей частью теряют эту способность. У круглых червей и коловраток клетки теряют способность к делению после прохождения эмбрионального развития, и рост организма, например у аскариды, происходит не за счет роста числа клеток, а за счет увеличения их размера.
В организме высших позвоночных клетки различных тканей и органов имеют неодинаковую способность к делению. Здесь встречаются клетки, полностью потерявшие свойство делиться: это большей частью специализированные, дифференцированные клетки (например, клетки центральной нервной системы). В организме есть постоянно обновляющиеся ткани (различные эпителии, кровь, клетки рыхлой и плотной соединительной ткани). В этом случае в таких тканях существует часть клеток, которые постоянно делятся (например, клетки базального слоя покровного эпителия, клетки крипт кишечника, кроветворные клетки костного мозга и селезенки), заменяя отработавшие или погибающие клеточные типы. Многие клетки, не размножающиеся в обычных условиях, приобретают вновь это свойство при процессах репаративной регенерации органов и тканей.
Примерно такие же типы клеток по способности их вступать в деление встречаются и у растительных организмов: это камбиальные клетки, дающие начало различным органам и тканям, клетки, интенсивно делящиеся, это клетки, возобновляющие деление при регенерации, это дифференцированные клетки, потерявшие в естественных условиях способность делиться. Клетки животных и растений, так же как одноклеточные эукарпотические организмы, вступают в процесс деления после ряда подготовительных процессов, важнейшим из которых является синтез ДНК.
Весь смысл клеточного деления заключается в равномерном распределении редуцированного генетического материала по двум новым клеткам.